|
OSMOSISLa osmosis es un caso especial de difusión en el que es el movimiento del disolvente el que se estudia, y se define en función de los solutos. Así, la osmosis es el movimiento del agua desde soluciones con baja concentración de soluto hasta soluciones con alta concentración de soluto.La osmosis puede ilustrarse (*) separando dos soluciones con concentraciones diferentes de soluto (sacarosa) por medio de una membrana semi-permeable (es decir una membrana que deja pasar el agua pero no la sucrosa. En este tipo de sistema, el agua pasara desde la solución A a la solución B, y este movimiento continuará hasta que se igualen las concentraciones (asumiendo que no hay cambios en el nivel hidrostático). Si este experimento se modifica de tal forma que la solución contenida dentro de la membrana semi-permeable tenga la misma composición que el líquido intracelular y la solución exterior es agua destilada, el agua atravesará la membrana haciendo subir el nivel del líquido en el tubo hasta 28 metros de altura. En este momento, el paso de moléculas de agua desde el exterior al interior se igualará y la presión de la columna de agua en el tubo contrarrestará el flujo desde A hasta B. Esta presión es la llamada presión osmótica y se expresa en mm de Hg o atmósferas. Como se ilustra en el ejemplo anterior, el líquido intracelular tiene una presión osmótica impresionante: 6.7 atmósferas. Sin embargo, dado que todos los fluídos del organismo tienen su propia presión osmótica, ninguna de las células está expuesta a presiones tan elevadas, sino que la presión osmótica en cada compartimento del cuerpo resulta de la diferencia de las respectivas presiones osmáticas. La presión osmótica, depende del número de partículas de la solución de acuerdo con la fórmula:
donde:
Dada la dificultad práctica debidas a la ionización y la formación de subgrupos osmóticamente activos en los fluídos biológicos, la presión osmótica se suele expresar en osmoles. Una solución conteniendo 1 mol de un soluto no ionizable en 1 litro de agua tiene una presión osmótica de 22.4 atm. Como resultado de esta definición, cualquier solución que tenga esta presión osmótica, independientemente de la concentración de sus solutos, se dice que tiene una presión de 1 Osm/litro. Como esta unidad es muy grande, en la práctica se utiliza el miliosmol (1 Osm = 1000 mOsm). La presión osmótica de los diferentes fluídos corporales está en los alrededores de 0.3 Osm o 300 mOsm y se determina experimentalmente a partir del punto de congelación. Como las membranas celulares son permeables al agua, usualmente no hay gradientes de presión osmótica entre compartimentos. Sin embargo, existen varios mecanismos locales que operan para producir pequeños pero significativas diferencias en las presiones osmóticas y que tienen importancia en: |
 |
EQUILIBRIO DE GIBBS-DONNAN (*) Cuando existen moléculas cargadas de gran tamaño que no difunden a través de una membrana semipermeable (como las proteínas), su presencia cambia la distribución de las partículas íonicas. En efecto, la proteína intracelular, cargada negativamente, atráe iones K+ y repele iones Cl-, produciéndose un gradiente eléctrico (simbolizado por las cargas + y - a ambos lados de la membrana) y sendos gradientes de concentración de K y Cl, iguales y de signo opuesto. En el equilibrio, se tiene: 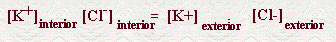
Debido a la naturaleza semi-permeable del endotelio capilar, las proteínas plasmáticas son retenidas en el compartimento vascular y su influencia sobre la actividad osmótica es capital para los movimientos de fluídos entre los compartimentos capilar e intersticial. El equilibrio de Gibbs-Donnan establecido a través del epitelio por la existencia de proteínas no difusibles añade un pequeño pero significativo incremento a esta actividad osmótica. Las proteínas del plasma originan una presión osmótica de unos 20 mm de Hg y la originada por las partículas cargadas producidas en el equilibrio de Gibbs-Donnan es de unos 6-7 mm de Hg. La suma de ambas es la presión oncótica o sea la atracción hacia el agua que ejercen las proteínas del plasma. TRANSPORTE ACTIVOCuando la producción de ATP de una célula es inhibida experimentalmente (por ejemplo por tratamiento de 2,4-dinitrofenol) la concentración de iones dentro de la célula se va aproximando gradualmente a la del exterior. Esto es debido a la lenta pérdida de iones a través de la membrana por los poros acuosos favorecida por el gradiente eléctrico y el gradiente de concentración. Eventualmente, la célula muere ya que sus enzimas citoplasmáticos requieren una baja concentración de Na+ y una alta concentración de K+. En todas las células, una cantidad significativa (hasta el 50%) de la energía producida, es decir de la energía almacenada en el ATP, se utiliza para mantener los gradientes iónicos de Na+, K+ y Ca++ a través de la membrana celular y a través de las membranas intercelulares.En al menos tres tipos de enzimas, la hidrólisis de ATP está directamente acoplada al transporte iónico.
|
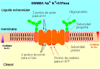 |
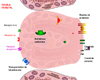
La Bomba Na+K+-ATPasaEl sistema de transporte Na+K+-ATPasa (*) ha sido estudiado muy detalladamente en células de riñón de mamífero. La proteína es un dímero, y cada subunidad contiene dos cadenas polipeptídicas diferentes. El proceso de transporte implica el paso de 3 iones Na hacia fuera de la célula y de 2 iones K hacia adentro por cada ruptura de una molécula de ATP.La proteína tiene en la parte orientada hacia el líquido extracelular dos puntos de fijación para el K+. La ouabaína (un potente glicósido cardíaco) inhibe la bomba Na+K+-ATPasa cuando se fija a una porción específica de la parte externa de la proteína, en donde usualmente se fijan los iones K+. La parte interna de la proteína tiene tres puntos donde se fijan los iones Na y un punto de fijación del ATP. Al fijarse el ATP a la proteína esta experimenta un cambio conformacional que impulsa los iones Na+ y K+ en contra de su gradiente de concentración. La Bomba Ca++-ATPasaOtra bomba importante en la función celular es la bomba Ca++-ATPasa que se encarga de mantener los niveles intracelulares de calcio a un nivel bajo. En algunas células, como los eritrocitos, está localizada en la membrana celular y su función es transportar Ca++ fuera de la célula. Sin embargo, en las células musculares, la bomba Ca++-ATPasa se encuentra en la membrana del retículo sarcoplásmico. La bomba transporta el Ca++ desde el citosol hacia el interior del orgánulo, que concentra y almacena el calcio. La salida del Ca++ del retículo sarcoplásmico al citosol muscular origina la contracción de la célula y se requiere una rápida eliminación de este calcio para que la célula se relaje.La bomba Ca++-ATPasa funciona de una forma muy parecida a la bomba Na+K+-ATPasa: dos iones de Ca++ son transportados fuera del citosol por cada molécula de ATP hidrolizado. La actividad de esta bomba está regulada de tal forma que si la concentración de Ca++ aumenta, la velocidad de bombeo aumenta hasta que la concentración citosólica se reduce a 0.1 mmolar. En el interior del retículo sarcoplásmico, existen dos proteínas capaces de acomplejar grandes cantidades de calcio: una de ellas, la calsecuestrina y Proteína fijadora del calcio con alta afinidad. Esta proteínas sirven como almacén de calcio y reducen las concentraciones de calcio libre en las vesículas sarcoplásmicas, reduciendo el gradiente en contra del cual tiene que trabajar la bomba. |
|
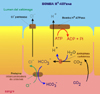
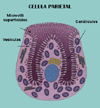
La Bomba de Protones (*) El jugo gástrico humano contiene una solución de ácido clorhídrico (H+Cl-) 0.1 M. Este medio fuertemente ácido desnaturaliza las proteínas ingeridas, facilitando la digestión por las enzimas proteolíticas, como la pepsina que funcionan a pH ácido. El ácido clorhídrico es producido por las células parietales (*) (o células oxínticas) que tapizan la mucosa gástrica. El proceso implica una ATPasa situada en la membrana apical de la célula, la parte que se enfrenta al lumen del estómago. El transporte activo de H+ hacia el exterior de la célula, ATP-dependiente, iría acompañado de un transporte pasivo de Cl- para mantener la electroneutralidad. Dicho transporte se haría a través de una Cl-permeasa. El resultado neto es que en el estómago hay una concentración de HCl un millón de veces mayor que en citoplasma de la célula. El protón excretado por la bomba de protones proviene de la hidratación del CO2 por una anhidrasa carbónica. El bicarbonato producido es, a su vez intercambiado por cloro en una proteína intercambiadora (*) ULTRAFILTRACIONLa ultrafiltración es el proceso mediante el cual las moléculas de un disolvente son obligadas a pasar a través de una membrana semipermeable en contra de un gradiente osmótico por efecto de una presión aplicada sobre la solución. El soluto es retenido a pesar de su elevada concentración.La ultrafiltración es un importante proceso fisiológico que juega un importante papel en la formación de orina y en las salidas de fluídos de los capilares. En ambos casos, la fuerza para vencer la presión osmótica es suministrada por la presión arterial. |
   |
|
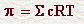
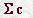 es la suma de las concentraciones de todas las partículas de soluto
es la suma de las concentraciones de todas las partículas de soluto