Introducci�nLa capacidad de las c�lulas eucariotas para adoptar diversas formas, en particular aquellas c�lulas especializadas como las c�lulas nerviosas con sus largos axones, las c�lulas epiteliales de la pared intestinal con sus microvilli, etc., y su capacidad para generar movimientos coordinados y dirigidos depende del citoesqueleto. El citoesqueleto es una compleja red de estructuras proteicas que se extiende por todo el citoplasma. Dado que el citoesqueleto est� ausente en las bacterias, puede haber sido un factor crucial en el desarrollo de las c�lulas eucar�ticas. Las diversas actividades del citoesqueleto dependen de tres tipos principales de filamentos constitu�dos por prote�nas:
Cada una de estas estructuras est� formada por diferentes mon�meros de prote�nas y puede adoptar diversas disposiciones espaciales seg�n la naturaleza de otras prote�nas asociadas. Algunas de estas, por ejemplo, unen los microfilamentos a la membrana, otras sirven para anclar las prote�nas de membrana y otras interaccionan con las primeras para crear movimientos, de los cuales los m�s estudiados son la contracci�n muscular y el movimiento ciliar. |
|
|
MICROTUBULOS: ESTRUCTURA Y FUNCIONLos microt�bulos est�n presentes en todas las c�lulas eucariotas desde las amebas hasta los animales, salvando contadas excepciones como los eritrocitos humanos. Son las fibras de mayor tama�o presentes en el citosol, con un di�metro aproximado de 24 nm y usualmente se agrupan formando haces en los que los diferentes t�bulos se unen mediante puentes para conferirles rigidez y solidez.
Funciones en las diferentes c�lulasLos haces de microt�bulos (*) adoptan diversas formas y aspectos y tienen funciones variadas:
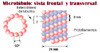
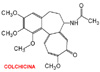
Muchos sistemas de microt�bulos constituyen estructuras l�biles, que pueden ser despolimerizados en determinadas circunstancias. Esta labilidad es esencial para su funci�n. Por ejemplo, la sustancia antimit�tica colchicina (*) despolimeriza los microt�bulos al unirse a la tubulina. Si se elimina la colchicina por lavado de la preparaci�n, se reconstituyen los microt�bulos. Composici�n de los microt�bulosTodos los microt�bulos est�n constitu�dos seg�n el mismo principio: el corte transversal de un microt�bulo (*) revela que est� constitu�do por 13 subunidades de 4 a 5 nm de di�metro con un hueco central. Los microt�bulos est�n compuestos por dos tipos de subunidades de prote�nas: la a-tubulina y la b-tubulina una de las cuales tiene un peso molecular de 55.000. La pared del microt�bulo est� formada por una matriz helicoidal de las dos subunidades. Hay 13 protofilamentos cada uno de ellos compuestos por d�meros que se extienden paralelamente al eje longitudinal del microt�bulo. Las unidades repetitivas son, pu�s, heterod�meros y est�n dispuestos dentro del microt�bulo alineados de cabeza a cola. Esto quiere decir que los microt�bulos tienen una polaridad definida y que los dos extremos no son estructuralmente equivalentesLos microt�bulos se forman por polimerizaci�n de los d�meros alfa-beta y, viceversa, cuando los microt�bulos se someten a diversas manipulaciones (p.ej congelaci�n) se disocian en sus respectivos d�meros. Por el contrario, los d�meros no se disocian en sus subunidades a menos de que se a�adan agentes desnaturalizantes. Asociados a las tubulinas, se encuentran adem�s una prote�nas minoritarias llamadas prote�nas asociadas a los microt�bulos (PAMTs) que desempe�an una funci�n estabilizadora e iniciadora de la reacci�n de polimerizaci�n. Diferentes estructuras microtubulares (husos mit�ticos o fibras axonales) contienen diferentes PAMTs Como se ha indicado anteriormente, los microt�bulos tienen una diferente polaridad porque sus subunidades de tubulina est� dispuestas seg�n una orientaci�n espec�fica en el pol�mero. Si se dejan polimerizar fragmentos purificados de tubulina sobre fragmentos de un axonema ciliar (-fragmentos que act�an como iniciadores o cebadores de la reacci�n de polimerizaci�n) se observa que la adici�n de d�meros en un extremo (llamado extremo A o extremo +) es varias veces m�s r�pida que en el otro extremo (llamado extremo D o extremo -). Igualmente, la despolimerizaci�n (o eliminaci�n de subunidades de cada uno de los extremos) tambi�n ocurre a diferente velocidad. En condiciones definidas de concentraci�n i�nica y de subunidades solubles, se llega a una condici�n de equilibrio (steady state) en la cual la adici�n de unidades de d�mero es compensada por la p�rdida de otras subunidades. En estas condiciones, como la adici�n tiene lugar preferentemente en la extremo A (+) y la p�rdida en el extremo D (-), el microt�bulo funciona como una "cinta transportadora". Un d�mero de tubulina incorporado en el extremo m�s viaja a lo largo del microt�bulo y eventualmente es despolimerizada en el extremo menos. |